2020年夏期ヤクシマザル調査報告
2020.9.27. 2020ヤクザル調査隊
1 はじめに
ヤクシマザル(Macaca fuscata yakui)は屋久島の亜熱帯性海岸林から標高1800mを越える山頂部まで連続的に分布することが知られている (丸橋他、1986; Hanya et al., 2004)。屋久島の西部海岸域では、1970年代から餌付けに頼らない長期にわたる継続調査が行われており、 多くの成果が得られている(Yamagiwa & Hill, 1998)。中高度・上部域に生息するヤクシマザルについては調査が遅れていたが、 1990年代に入って、西部海岸部以外での調査が本格化し、上部域のニホンザルの密度、群れサイズ、食性、活動時間配分など、 基本的な生態学的情報が、すこしずつ明らかになってきた(Hanya et al., 2003a; Hanya, 2004a, 2004b)
「ヤクザル調査隊」は1989年以降、毎年夏にヤクシマザルの分布調査を行ってきた。このうち、1994、1995、1997年には、 自然植生の垂直分布が連続して保存されている西部域で調査を行い、以下のことが明らかになった。 (1)標高300mまでの海岸部には、1km2あたり約4群と、きわめて高密度で分布している。(2)標高300mから800mの照葉樹林帯、 標高800mから1,200mの照葉樹林・ヤクスギ林移行帯、標高1200m以上のヤクスギ帯では、群れ密度は1km2あたり 1.2群から1.6群と、ほぼ一定である。(3)群れサイズについては正確な情報は得られなかったが、標高800m以上の移行帯およびヤクスギ帯では、 それ以下の標高帯に比べ小さいようであった(Yoshihiro et al., 1999; Hanya et al., 2004)。
さらに1998年からは、西部域の大川林道終点地域で継続調査を行うことにした。具体的には、調査域内の群れの正確な分布と、 対象とした群れのサイズと性年齢構成を明らかにすることを目的として行われた。これらの資料を蓄積することによって、屋久島中高度域 におけるヤクシマザルの個体群動態のメカニズムを明らかにすることが最終的な目的である。1998年の調査では、大川林道終点地域の、 瀬切川の両岸で調査を行い、1群の構成を確定したほか、3群について、大まかな構成を確認した。群れサイズはどれも20頭を越えるものではなかった。 2000年には、HR群を全頭を個体識別し、ほかにもPE、OM、SY群の合計4群が識別できるようになった。2001年の調査では、 さらにBR群が識別されるようになった。BR群は2003年以降は調査域内で観察されなくなったが、2005年にはSY群がSS群、 YY群の二つの群れに分裂し、現在調査域内では5群を識別している。これらの群れの子持ち率(または粗出産率、 観察されたオトナ・ワカモノメスのうち、アカンボウを持っていたメスの割合)は年によって4%-43%のあいだで変動した。 2000年から、これら人口学的資料の収集と並行して、ブロック分割定点調査法による集団密度の推定を行った。 この地域は自然林と伐採地が混在しており、しかも伐採された年数が異なっている。このような地域で集団密度の推定を行うことで、 ニホンザルが植生の撹乱の程度に応じてどのように土地利用のやり方を変化させていくのかを明らかにし、さらにその調査を毎年継続して行うことで、 伐採後の植生の変化に応じて土地利用のあり方がどう変化していくかを明らかにして行くことが目的である。2000年から2003年までの結果については、Hanya et al. (2005)に発表した。
さらに2003年度からは、定点調査中にヒトリザルおよびシカについてもサルの集団と並行して資料を収集するようにした。 シカ調査は、2008年までは定点で音声が聞こえる頻度を記録していたが、この方法で得られた結果の信頼性に疑問があったため、2008年度からは、糞塊法による調査を開始した。 定点の周辺にシカ糞調査用の調査区を設置し、シカ密度の年変動を恒久的に調査できるようになった。サルの分布と関連すると思われる、 森林の果実量調査は、自然林では1999年から、伐採地では2002年から継続して行っている。また、2010年から、定点でのヒルの調査も行った。 これらの結果の概要についても、この報告書で報告する。
2 調査地域と植生
調査地は、屋久島西部の瀬切川上流の地域である(図1)。 標高は750mから1350mにあたる。調査地域の面積は7.5km2である。
調査域は全体として照葉樹林・ヤクスギ林移行帯にあたる。イスノキ、ウラジロガシなどの照葉樹が、スギ・モミ・ツガなどの針葉樹と混交している。 林床はハイノキを中心とする常緑低木に広く覆われている。調査域は大川林道を中心として植生の撹乱があり、伐採跡はスギの幼樹、ヒサカキやハイノキなどの低木に覆われている。 伐採後の年数は大川林道の終点近くや12支線沿線では22-33年程度で、西(入口方向)へ行くほど年数が経っている。もっとも西側の5班の調査域付近ではおよそ40年程度である。 伐採後の更新方法には2種類ある。一つは伐採後自然の更新過程に任せる天然更新で、12支線沿いや大川林道の終点付近からおおむね3班のdf付近まではこの方法で行なわれている。 もう一つは伐採後スギを植林する人工更新で、それより西の、3cdおよび4班と5班の調査域付近で行なわれている。
3 調査方法
参加者は、11人である。今年は、新型コロナウイルス感染防止対策として、調査の規模を大幅に縮小し、各自が参加できる期間に参加することとした。
サルの集団の調査
例年行っているブロック分割定点調査による調査は行わなかった。対象となる5つの群れ(HR群、PE群、OM群、SS群、YY群)を探し、可能な限り追跡して、行動圏と性年齢構成を調査した。
ヒトリザル
実施しなかった。
シカ調査
サル調査の定点近傍に、4メートル×50メートルのシカ調査区画を設け、その中に落ちているシカの糞塊数を、それぞれの糞塊の中の糞粒数とともに記録した。 この調査は、統括者が交替で行った。また、糞はすべて採取して、ここの糞粒の長径と短径を計測した。
ヒル調査
生きたヒルを採取した。吸血していなかった約30個体は、三重県いなべ市の「こどもヤマビル研究会」に提供し、吸血していた9個体は、生活史の調査のため、愛知県犬山市の霊長類研究所まで持ち帰って飼育した。
果実量調査
これまでの調査で、5メートル四方の植生調査区を、伐採地に26個、自然林に10個設置してある。 その中に生えている、サルが食べる液果をつける樹木(ヒサカキ、ハイノキ、オニクロキ、サカキ)について、結実数を直接計数した。
5 調査日程
(かっこの中はその日離島した人を除く調査員の数)
8/5 集合(5)
8/6 入山、シカ調査
8/7 シカ調査(5)
8/8 シカ調査、下山(5)
8/9 日帰りで果実量調査、OM群調査(6)
8/10 休憩(5)
8/11 入山、果実量調査、OM群調査(5)
8/12 果実量調査、OM群調査(3)
8/13 果実量調査、OM群調査、下山(4)
8/14 日帰りでOM群(4)
8/15 休憩(2)
8/16 入山、OM群、SS群調査(2)
8/17 OM群、SS群調査(2)
8/18 OM群、SS群調査(2)
8/19 SS群調査(3)
8/20 果実量調査、OM群、SS群調査(3)
8/21 HR群、OM群、PE群調査(3)
8/22 HR群、PE群調査、下山(3)
8/23 休息(4)
8/24 日帰りでHR群、PE群調査(3)
8/25 日帰りでYY群調査(3)
8/26 台風のため待機(3)
8/27 台風のため待機(3)
8/28 日帰りでYY群調査(4)
8/29 日帰りでHR群、PE群調査(5)
8/30 日帰りでYY群調査(4)
8/31 日帰りでYY群調査(4)
9/1 台風のため待機(4)
9/2 台風のため待機(4)
9/3 日帰りでYY群調査(4)
9/4 解散
6 調査結果
長時間追跡できた集団の遊動図を図3に、それらをもとにして識別できた群れの遊動図を図4に示した。また、群れのカウント例の一覧と、それをもとにして分かった、重複を除いた最低の群れサイズ、性年齢構成を表1に、識別できた個体の記録を図5に挙げた。
以下に、調査域全体のシカの分布、果実生産の傾向、調査した5つの群れの遊動域と遊動パターン、サイズと構成などの情報について述べる。
シカ糞塊数
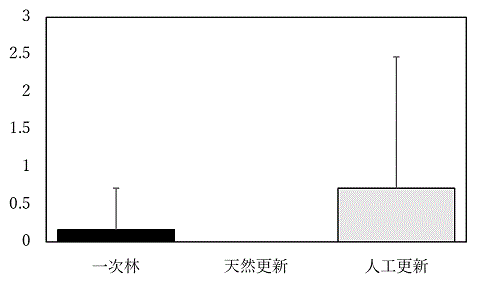
トランセクトあたり(50m*4m)の平均糞塊数の95%信頼区間は0.2±0.3である。最大値は5b(人工更新)の5、最小値は0であった。調査地では2010年4月から2011年3月にかけて、シカの捕獲が行われていた。 糞塊数は2009年は3.1±1.34、2010年は1.2±0.58、2011年は1.1±0.52、2012年は2.5±1.1、2013年は4.50±1.81、2014年は1.60±0.78、2015年は1.00±0.47、2016年は0.6±0.46、2017年は1.2±2.57、2018年は0.7±1.05、2019年は0.8±0.40だった。 糞塊数の植生による違いは、すべての組み合わせで有意でなかった(Tukey-Krammer法)。
図A シカ糞塊密度の植生による変異。y軸の単位は個/50m*4m。平均+標準偏差を示す。
果実量
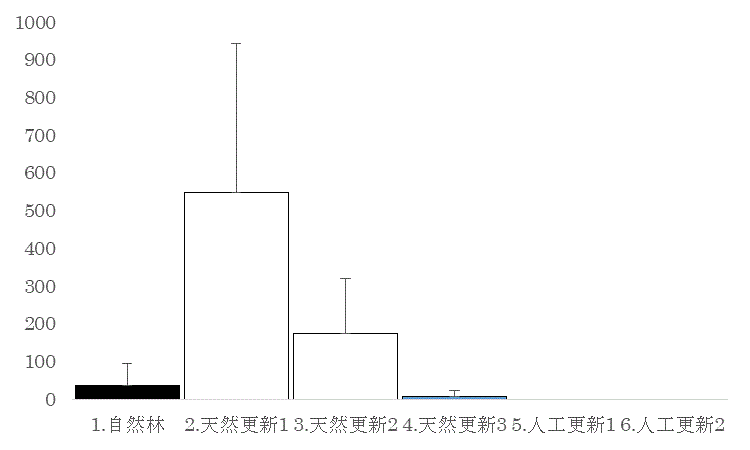
5m四方の調査区内で結実していた液果の果肉重量の植生による変異を図Bに示した。果実生産量は天然更新1で最大で、天然更新2、自然林、天然更新3がそれに次ぐ。 人工更新地では果実生産が見られなかった。これは、過去の年の調査と同様の傾向である。 天然更新地1と天然更新2、天然更新3、人工更新1、人工更新2、自然林の間の組み合わせで、有意な差があった (Tukey-Krammer法、p<0.05)。
図B 果実生産量の植生による違い。平均+標準偏差を示す。単位は果肉部分の湿重で、g/25m2を示す。
HR群
HR群はオトナを中心にほぼ全頭が個体識別されている群れである。今年の調査では、3日間追跡を行った。 東は1d定点付近、西は12支線終点道上の1161メートルピーク付近までを利用することが確認された。 行動圏の多くは、伐採されていない。2020年3月・4月に構成を確認しており、それに今回確認したアカンボウを加えると、オトナオス5頭、オトナメス8頭、コドモ8頭、アカンボウ4頭の合計25頭だった。 昨年生まれた2頭のアカンボウのうち、1頭は確認できたが、もう1頭は確認できなかった。
PE群
PE群は、西は2fから2h定点に向かう尾根の西から、東は1e・2f中央の尾根付近、南は2h定点付近までの非常に狭い範囲でのみ発見された。 ただし、2h定点の南から声が聞こえることがあったので、おそらく調査域よりもさらに南に広がっていると考えられる。 12支線の道上と道下の両方を利用していたため、PE群カウントを目的とした5日で毎日出現が確認された。特に8月29日はほとんどの個体が道上に移動したため、多数の個体のカウントができた。 確認された個体は、オトナオス2頭、オトナメス3頭、コドモ5頭、アカンボウ2頭の合計12頭だった。 昨年生まれていたアカンボウ1頭の生存は確認できなかった。
OM群
OM群は、大川林側本線上の中村街道入口付近から3d定点西の谷付近で観察され、また12支線側の中村街道入口の東の谷から大曲より南西に位置する尾根で観察された。 今年度OM群が遊動した東端は2c定点の西側谷で、西端は3dの西側の谷であった。 一方で北端は不明であるが、少なくとも2cと3d定点間の北側を遊動しているようだ。南端は大曲南西の尾根よりやや南だと思われる。 今年度OM群は移動中に偶然出会ってカウントした調査日も含めると、計8日間調査された。 構成はオトナオス3頭、オトナメス4頭、コドモ5頭、アカンボウ1頭の計13頭であった。昨年アカンボウは確認されておらず、今年も1歳は確認されなかった。
OM群とHR群の行動圏の境界付近に不明な群が出現した。具体的には大川林道本線側の中村街道入口周辺と12支線2e定点の北側入口の西側の道上で出現した。 後者の際に観察者(半谷、本田)の2名でHR群でもOM群でもないことを確認した。 群れのオスで人にかなり慣れている個体がいたが、観察者を見ると逃げる個体もいた。
SS群
SS群は大川林道上の辻南から東の谷付近から4d定点よりやや西付近で観察された。 4d定点付近からは4f定点の方向へ移動していた。これらはSS群の行動圏の東端西端と一致する。 北端は3b定点付近まで、南端は追跡できなかったため不明だが、辻南から4fの間の南側までは利用していた。 SS群は5日の調査中すべての日でカウントでき、構成はオトナオス3頭、オトナメス8頭、コドモ5頭、アカンボウ4頭の計20頭であった。 昨年確認されていたアカンボウ1頭の生存は確認できた。
YY群
YY群の行動圏は、大川林道の4班と5班の境界付近が東の端で、大川林道の瀬切川・大川の分水嶺付近(万里の長城の南)を西の端である。 今年の調査では、大川林道の分水嶺よりもさらに入り口側、P832の南東の林道がくびれている部分にも出現することが確認された。 これは、今年に限らず、過去の調査でもっとも一番南にYY群が出現した記録である。 今年は5日間追跡調査を行った。そのうち8月25日だけは4班に近い場所に出現したが、そのほかの日は5班領域に出現し、行動圏の中でも西側を中心に利用していた。 確認された個体は、オトナオス5頭、オトナメス6頭、コドモ5頭、アカンボウ3頭の、合計19頭だった。 9月3日に万里の長城の南で不明群が現れた。
謝辞
この調査を行うにあたって、屋久島森林生態系保全センターには、調査を許可していただきました。調査の準備段階で、多くの調査隊OB、OGの方にも御支援を頂きました。 厚くお礼を申し上げます。
引用文献
Buckland ST, Anderson DR, Burnham KP & Laake JL (1993) Distance Sampling: Estimating Abundance of Biological Populations. Chapman and Hall, London.
Hanya G, Noma N & Agetsuma N (2003a) Altitudinal and seasonal variations in the diet of Japanese macaques in Yakushima. Primates 44: 51-59.
Hanya G, Yoshihiro S, Zamma K, Kubo R & Takahata Y (2003b) New method to census primate groups: estimating group density of Japanese macaques by point census. American Journal of Primatology 60: 43-56.
Hanya G (2004a). Diet of a Japanese macaque troop in the coniferous forest of Yakushima. International Journal of Primatology 25: 55-71.
Hanya G (2004b). Seasonal variations in the activity budget of Japanese macaques in the coniferous forest of Yakushima: effects of food and temperature. American Journal of Primatology 63: 165-177.
Hanya G, Yoshihiro S, Zamma K, Matsubara H, Ohtake M, Kubo R, Noma N, Agetsuma N & Takahata Y (2004). Environmental determinants of the altitudinal variations in relative group densities of the Japanese macaques on Yakushima. Ecological Research 19: 485-493.
Hanya G, Zamma K, Hayaishi S, Yoshihiro S, Tsuriya Y, Sugaya S, Kanaoka MM, Hayakawa S & Takahata Y (2005) Comparisons of food availability and density of Japanese macaques in primary, naturally regenerated and plantation forests. American Journal of Primatology 66: 245-262.
丸橋珠樹,山極寿一,古市剛史 1986. 屋久島の野生ニホンザル. 東海大学出版会.
Yamagiwa J and Hill DA (1998). Intraspecific variation in the social organization of Japanese macaques: past and present scope of field studies in natural habitants. Primates, 39: 257-273.
Yoshihiro S, Ohtake M, Matsubara H, Zamma K, Hanya G, Tanimura Y, Kubota H, Kubo R, Arakane T, Hirata T, Furukawa M, Sato A and Takahata Y (1999). Vertical distribution of wild Yakushima macaques (Macaca fuscata yakui) in the western area of Yakushima island, Japan: preliminary report. Primates 40: 409-415.